Der weltweite Austausch von Genetik und die daraus folgende Konzentration auf immer weniger Rassen in der Milchproduktion hat zur Folge, dass immer häufiger die gleichen Kuhtypen in den unterschiedlichsten Umwelten gehalten werden. Darum stellt sich die Frage, ob der genetische Teil einer Leistung (Milch, Gesundheit, Fruchtbarkeit etc.) einer Kuh nicht auch von der Umwelt abhängig ist.
Das würde heissen, dass in den verschiedenen Umwelten unterschiedliche Gene oder ein unterschiedliches Zusammenspiel der Gene für die Ausprägung der Merkmale verantwortlich sind. Sollte dies der Fall sein, spricht man von Genotyp x Umwelt (GxE) Interaktionen. GxE Interaktionen können ganz verschieden ausgeprägt sein. Wir möchten dies anhand der nachfolgenden Beispiele in Tabelle 1 kurz erläutern.

Foto: swissherdbook
Rangierung von fiktiven Stieren in unterschiedlichen Umwelten
| Stier | Talzonen | Bergzonen | Erklärung, bzgl. GxE Interaktionen | ||
|---|---|---|---|---|---|
 |
 |
||||
| ZW | Rang | ZW | Rang | ||
| Gino | 122 | 1 | 120 | 1 | Keine GxE Interaktionen; Rangierung und Abstände zwischen den ZW ist gleich. |
| Turo | 105 | 2 | 103 | 2 | |
| Leno | 97 | 3 | 95 | 3 | |
| Robi | 89 | 4 | 87 | 4 | |
| Gino | 122 | 1 | 122 | 1 | Keine GxE Interaktionen; Rangierung ist gleich, die Abstände zw. den ZW verändern sich. Man spricht von Skalierungseffekten. |
| Turo | 105 | 2 | 101 | 2 | |
| Leno | 97 | 3 | 90 | 3 | |
| Robi | 89 | 4 | 80 | 4 | |
| Gino | 122 | 1 | 120 | 1 | Leichte GxE Interaktionen; Rangierung und Abstände zw. den ZW sind leicht verändert. |
| Turo | 105 | 2 | 94 | 3 | |
| Leno | 97 | 3 | 98 | 2 | |
| Robi | 89 | 4 | 89 | 4 | |
| Gino | 122 | 1 | 90 | 4 | Starke GxE Interaktionen; Rangierung und Abstände zw. den ZW sind stark verändert |
| Turo | 105 | 2 | 98 | 3 | |
| Leno | 97 | 3 | 118 | 1 | |
| Robi | 89 | 4 | 112 | 2 | |
Tabelle 1: Rangierung von fiktiven Stieren anhand ihrer Zellzahl-Zuchtwerte (ZW) in zwei unterschiedlichen Umwelten (Talzone und Bergzone).
Das klassische Verfahren, um das Vorhandensein von GxE Interaktionen zu prüfen, ist die Anwendung von Zwei- oder Mehrmerkmalsmodellen. Falconer (1952) hat die Grundsätze dieses Ansatzes bereits vor langer Zeit beschrieben. Heute wird dieses Modell wie folgt angewendet: Ein Merkmal wird entsprechend der Anzahl Umwelten, in denen es beobachtet werden kann, aufgeteilt. Anschliessend werden die genetischen Korrelationen (ra) zwischen diesen gesplitteten Merkmalen berechnet. Wenn ra kleiner als 0.8 ist, spricht man von stärkeren GxE Interaktionen (Robertson, 1959 und Boelling, 2003). Wenn dies der Fall ist, muss man sich überlegen, ob Zuchtprogramme aufgeteilt werden müssen: Zuchtwerte, die in einer Umwelt geschätzt werden, hätten nur noch begrenzte Gültigkeit in anderen Umwelten. Dies macht deutlich, dass Leistungsprüfungen möglichst in den Umwelten stattfinden sollen, in denen auch die tatsächliche Produktion geschieht. Zusätzlich unterstreicht dies die Notwendigkeit von Interbull, damit Zuchtwerte international vergleichbar gemacht werden können.

In dem Projekt „2-Org-Cows“, in welchem die Eignung von Zweinutzungsrassen für Bio- und Graslandbasierte Produktionssysteme untersucht wurden, hat die Qualitas AG umfangreiche Analysen zum Thema GxE Interaktionen durchgeführt.
Vor allem sind wir der Frage nachgegangen, ob spezialisierte Milchrassen anders reagieren als Zweinutzungsrassen. In einem ersten Schritt haben wir die oben beschriebenen Zweimerkmalsmodelle angewendet. Die wichtigsten Resultate sind in den nachfolgenden Tabellen 2 und 3 zusammengestellt.
Genetische Korrelationen der Umwelten Berg- und Talzonen
| Umwelten | Bergzone(n) | ||
| Merkmale | Milch kg | Zellzahlen | |
| Talzonen | Milch kg | 0.93, 0.94, 0.92 0.95, 0.93 |
|
| Zellzahlen | 0.90, 0.90, 0.91 0.94, 0.95 |
||
Tabelle 2: Genetische Korrelationen zwischen den aufgeteilten Merkmalen Milch kg und Zellzahlen (jeweils Laktationsmodell) auf die Umwelten Berg- und Talzonen. Rassen: SI, SF, RH/HO, OB/ROB, BV/BS
Genetische Korrelationen der Umwelten biologische Landwirtschaft und konventionelle Landwirtschaft
| Umwelten | Konventionelle Landwirtschaft | ||
| Merkmale | Milch kg | Zellzahlen | |
| Bio. Landwirt. |
Milch kg | 0.99, 0.98 | |
| Zellzahlen | 0.97, 0.97 | ||
Tabelle 3: Genetische Korrelationen zwischen den aufgeteilten Merkmalen Milch kg und Zellzahlen (jeweils Laktationsmodell) auf die Umwelten biologische Landwirtschaft und konventionelle Landwirtschaft. Rassen: OB/ROB, BV/BS
Die Zahlen zeigen, dass in den untersuchten Umwelten in der Schweiz wenig bis keine GxE Interaktionen vorliegen. Demzufolge sind Anpassungen in den Zuchtprogrammen nicht notwendig. Ähnliche Untersuchungen aus anderen Ländern, zum Beispiel aus Österreich, bestätigen diese Resultate (Pfeiffer et al., 2016).

Es muss aber vermutet werden, dass eine kategoriale Aufteilung der Umwelten, wie in der vorliegenden Analyse angewendet, nicht geeignet ist, um die Interaktionen zwischen den Genotypen und einer spezifischen Umwelt beschreiben zu können. Darum werden immer mehr sogenannte Reaction Norm Models (RNM) angewendet (z.B. Falconer und Mackay, 1996; Calus et al., 2002; Calus und Veerkamp, 2003). Dabei versucht man die Reaktion der Genotypen auf kontinuierlich verändernde Umweltbedingungen zu analysieren. Das klassische Beispiel ist das Hinzufügen eines kontinuierlichen Umwelteffektes in ein Testtagesmodel.
Im Rahmen des Projektes „2-Org-Cows“ wurde, in Zusammenarbeit mit der Universität Giessen, jedem Testtag über einen Zeitraum von zehn Jahren ein Index aus Temperatur und Luftfeuchtigkeit zugeordnet und als weiterer Effekt im Testtagesmodell berücksichtigt (THI: Temperatur humidity index (NRC, 1971)).
Am Beispiel der Rassen Red Holstein/Holstein und Simmental kann ein klarer Einfluss des THI auf die phänotypischen Leistungen gezeigt werden. Abbildung 1 zeigt, dass ab einem THI von ungefähr 50 die täglichen Proteinleistungen abnehmen, beziehungsweise die Zellzahlen pro ml Milch zunehmen. Bei den Rassen Braunvieh und Original Braunvieh ist zu sehen, dass die Verläufe der Kurven beinahe identisch sind.
Auswertungen der Varianz/Covarianz-Struktur aus dem RNM entlang des THI und der Tage in Milch (DIM) bestätigen, dass sich die Varianz entlang des Umweltgradienten (THI) verändert. Dies ist in Abbildung zwei dargestellt. Es zeigt sich aber auch, dass die Veränderungen entlang DIM eher grösser sind. Es bestehen Unterschiede zwischen Braunvieh und Original Braunvieh.
Diese Resultate zeigen, dass für jede THI x DIM Kombination für alle Tiere Zuchtwerte geschätzt und so Tiere für spezifische Umweltverhältnisse selektiert werden könnten. Die Analysen brachten ebenfalls zutage, dass die Zusatzinformationen die Schätzverfahren etwas sicherer machen. Realistischerweise müssten aber der Aufwand für die zusätzliche Datenbeschaffung und längere Rechenzeiten sowie der Ertrag, also höhere B%, einander gegenübergestellt werden, um abschätzen zu können, ob sich dieser Schritt im Routinebetrieb lohnen würde.

Was aber weiterzuverfolgen ist, sind die individuellen Zuchtwert-Verläufe der Kühe und Stiere entlang der DIM x THI Kombinationen. Tiere, die weniger stark mit Zuchtwert-Schwankungen auf äussere Einflüsse reagieren, sind heutzutage gewünscht. Das wäre eine Möglichkeit, wie man resiliente Tiere züchten könnte. Genaue Validierungen mit Gesundheits- und Fruchtbarkeitsdaten sind diesbezüglich notwendig. Zusätzlich muss die Frage beantwortet werden, ob der THI als Platzhalter für andere kontinuierliche Umwelten (z.B. Fütterung, Management) verwendet werden kann? Eine Beispielfrage könnte sein, ob eine Kuh, die auf hohe THI Werte reagiert, auch mit einer schlechten Fütterung zurecht kommt?
Die Qualitas AG sieht in diesem Bereich grosses Potential: mit der immer besseren und hochauflösenden Datenerfassung durch Livestock precision farming, zum Beispiel durch Roboter- und Sensordaten, oder Meteo- und Futterdaten, entsteht eine grössere Datendichte. Diese erlaubt es, genetische Analysen durchzuführen, um resiliente Tiere identifizieren und selektieren zu können.

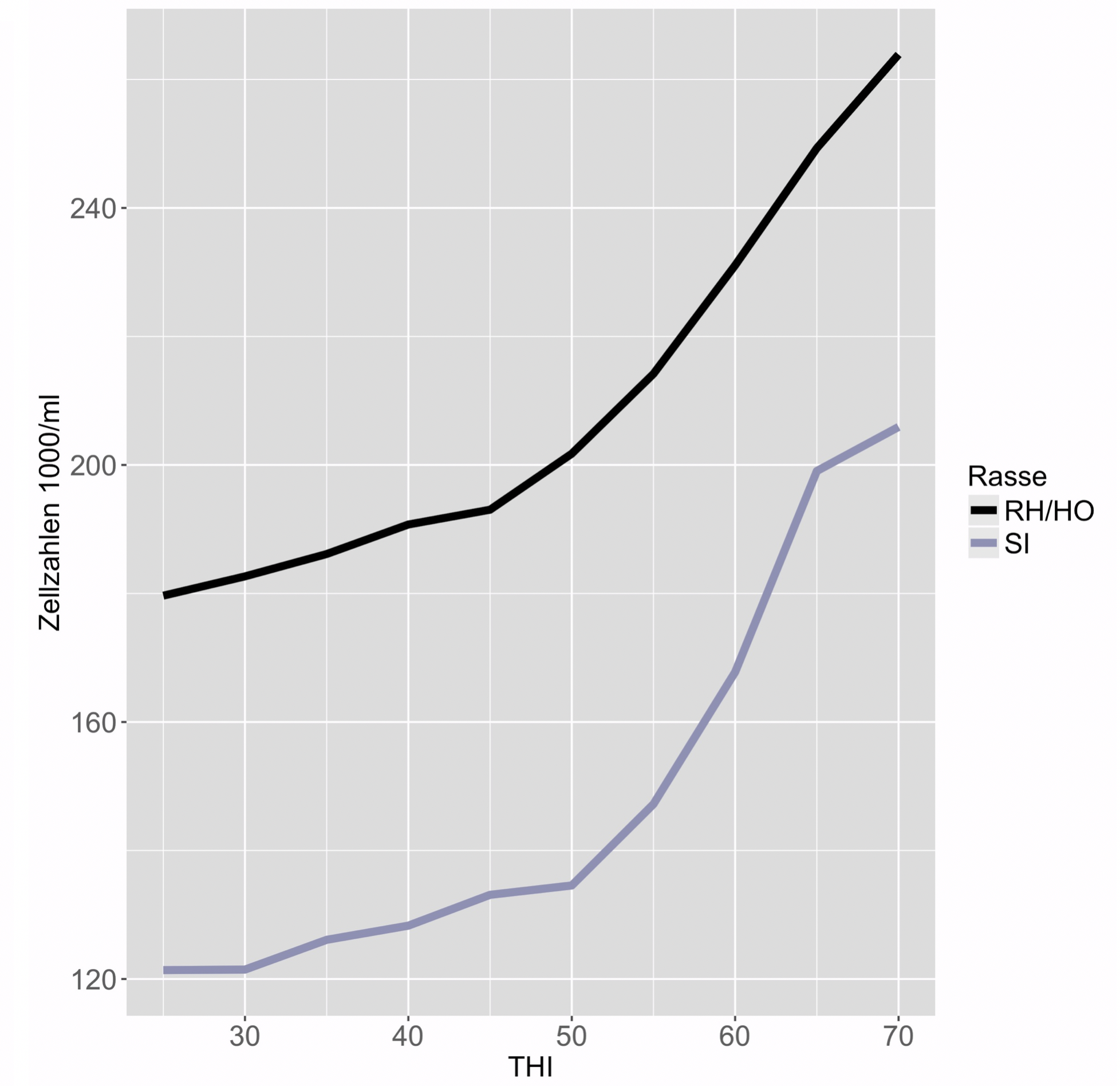
Abbildung 1: Testtagesleistungen (Protein und Zellzahlen) in Abhängigkeit des THI von den Rassen Red Holstein/Holstein (RH/HO) und Simmental (SI) in den ersten 100 Tagen der Laktation (1-100 DIM), für die Laktationen 3 und folgende (3ff).

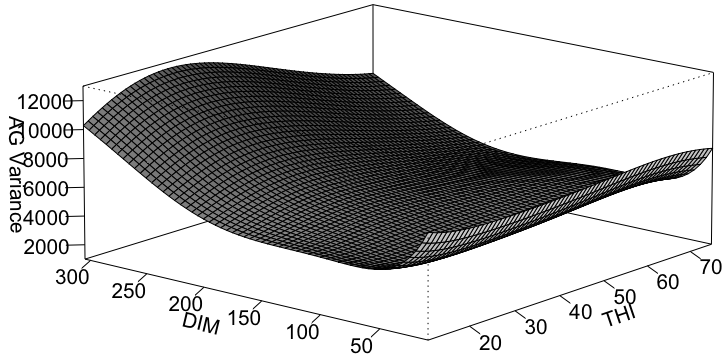
Abbildung 2: Additiv genetische Varianz entlang DIM und THI. Aufgeteilt auf Braunvieh (rechte Grafik) und Original Braunvieh (linke Grafik).
Literatur:
- Boelling D, Groen AF, Sørensen P, Madsen P and Jensen J (2003) Genetic improvement of livestock for organic farming systems. Livest Pro Sci 80:79–88
- Calus MPL, Groen AF, De Jong G (2002) Genotype × environment interaction for protein yield in Dutch dairy cattle as quantified by different models. J Dairy Sci 85:3115–3123
- Calus MPL, Veerkamp RF (2003) Estimation of environmental sensi- tivity of genetic merit for milk production traits using a random regression model. J Dairy Sci 86:3756–3764
- Falconer DS (1952) The problem of environment and selection. Am Nat 86:293-298
- Falconer DS, Mackay TFC (1996) Introduction to quantitative genetics, 4th edn. Pearson Education Limited, England
- NRC (1971) A Guide to Environmental Research on Animals. Natl Acad Sci, Washington, DC
- Robertson A (1959) The sampling variance of the genetic correlation coefficient. Biometrics 15:469–485
- Pfeiffer C, Fuerst C, Schwarzenbacher H and Fuerst-Waltl B (2016) Genotype by environment interaction in organic and conventional production systems and their consequences for breeding objectives in Austrian Fleckvieh cattle. Livest Sci 185:50–55
Beitragsfoto auf Übersichtsseite: Braunvieh Schweiz

